Tema 6: Semilla
|
|
|
6.8. Embrión y sustancias de reserva
El embrión es una plantita en miniatura en estado de vida latente o letargo.
Se
forma generalmente como consecuencia de la fecundación
de la ovocélula.
La doble fecundación en Angiospermas da lugar al
desarrollo del embrión y del endosperma, tejido
nutricio.
|
Ciclo
de vida de las Angiospermas

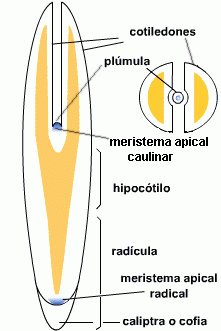
El embrión está formado por la
radícula dirigida hacia la micrópila, el hipocótilo que es
el corto eje caulinar, los cotiledones que son
las primeras hojas y la
plúmula o gémula que es el ápice caulinar y a veces algunos primordios
foliares.
En las Pteridófitas el embrión es unipolar:
sólo se
establece el polo de crecimiento correspondiente al
vástago, pues las raíces son adventicias. En cambio en las plantas con
semilla, el embrión es bipolar: en un polo se
desarrolla el vástago, y en el otro la raíz.
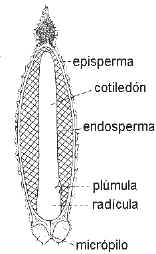
En las
Gimnospermas el embrión presenta generalmente dos
a varios cotiledones, dos en Ginkgo
y 5-18 en Pinus.
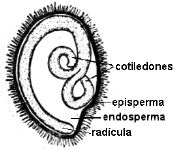
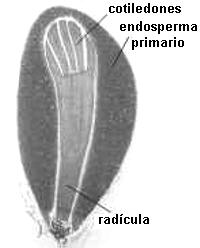
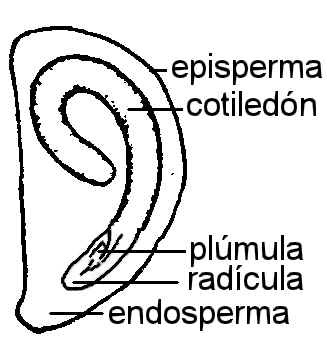
En Dicotiledóneas presenta dos cotiledones que pueden tener
diverso aspecto, foliáceos como en el zapallo y ricino; carnosos como en el
maní y arveja; con los extremos retorcidos como en el tomate, plegados de
diversas maneras, características para cada género o familia.
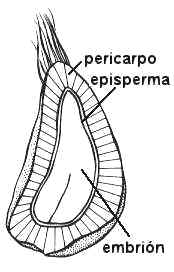
| fig. 6.81. Embrión de Ricinus
communis en corte longitudinal de semilla |
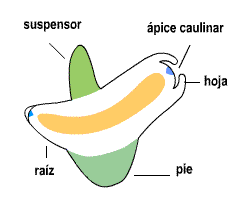
Fig. 6.82. Embrión
de Phaseolus sp., poroto |
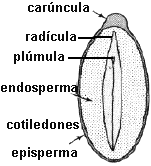
Fig. 6.83. Embrión de Lycopersicum
esculentum, |
 |
 |
 |
| Esquema de
Esau (1977) |
Esquema de
Fahn (1989) |
Esquema de
Hayward (1953) |
En
Monocotiledóneas presenta un solo cotiledón: su posición es lateral, igual
que la de la plúmula. En Allium es cilíndrico.

|
fig. 6.84. Embrión de Pinus
taeda en corte longitudinal de semilla |
Fig. 6.85. Embrión de Allium
cepa, cebolla, en corte longitudinal de semilla |
fig. 6.86. Embrión de Typha angustifolia,
en corte
long. de semilla albuminada
|
|
|
 |
|
Foto de Stone &
Gifford (1997) |
Esquema de Hayward
(1953) |
Esquema de Goebel
(1933) |
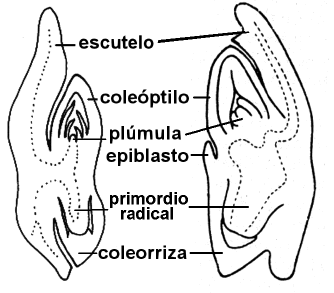
En Gramíneas el embrión completamente
desarrollado es bastante complejo: presenta las siguientes
partes:
 |
Escutelo: cotiledón transformado en órgano
absorbente, adosado al
endosperma. La epidermis abaxial es un epitelio secretor, segrega enzimas que
solubilizan las sustancias de reserva, las absorbe y las transporta al embrión.
En
algunas especies de Avena y otros géneros el ápice del escutelo se alarga e
invade el endosperma; superficialmente presenta papilas. |
|
Plúmula:
presenta varios primordios foliares; en el embrión del trigo están
presentes 6 de las 10 hojas que desarrolla la planta en toda su vida. |
|
Coleóptilo:
es una vaina cerrada que encierra la plúmula. Presenta, en el momento de la
germinación, un orificio apical por donde saldrá la plúmula. Según la
interpretación más aceptada es la primera hoja, ya que Streptochaeta y Jouvea
pilosa, gramíneas primitivas, presentan el coleóptilo abierto, con un haz
mediano y márgenes libres. |
|
Coleorriza:
es la vaina que envuelve la radícula y la
caliptra. En embriones jóvenes se continúa con el suspensor.
Se interpreta como la raíz primaria
abortiva o degenerada, y es perforada por la radícula en
el momento de la germinación. Excepcionalmente, especies de otras familias también presentan esta estructura:
Commelinaceae (monocotiledóneas), Cycadaceae (gimnospermas) y
Lauraceae (dicotiledóneas). |
|
Primordio radical:
interpretado como
la primera raíz adventicia, considerando que la coleorriza
es la raíz primaria. |
|
Epiblasto: es un
apéndice opuesto al
escutelo, que falta en
algunas gramíneas. Es un órgano escuamiforme, que según la interpretación de
distintos investigadores sería un cotiledón vestigial, o una extensión de la
coleorriza, o la vaina del cotiledón.
El procámbium se extiende como un
cordón en el embrión. Permite reconocer el nudo escutelar, inmediatamente
encima de la radícula, consecuentemente no hay hipocótilo. Las raíces
adventicias nacen por encima de este nudo. |
|
Fig. 6.87. Embriones
de gramíneas, Zea (maíz) y Triticum (trigo) |
 |
Hay
casos especiales de desarrollo del embrión: en las Orchidaceae
el embrión es totalmente indiferenciado; en Cuscuta, planta parásita,
el embrión sólo presenta plúmula, carece de cotiledones y de radícula.
|
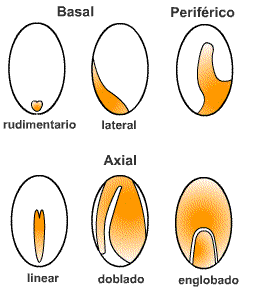
Fig. 6.88. Clasificación de los
embriones |
|
Martin (1946)
hizo una clasificación de la
semilla basada en la posición, el tamaño y la forma del embrión. Las
categorías básicas son embrión basal, embrión periférico
y embrión axial.
Los basales, según el
tamaño, se clasifican en rudimentario, amplio, capitado y lateral (gramíneas).
Los axiales son los más frecuentes, y hay varios tipos según forma y tamaño:
lineal, pigmeo, micro, espatulado, doblado, plegado y englobado. |
 |
Las semillas raramente carecen de sustancias de
reserva; las mismas están ausentes en las semillas de Orchidaceae.
En Hymenocallis (Liliaceae) los
tegumentos son verdes y con estomas; el desarrollo embrionario está en
conexión con la actividad de este tejido.
En las Gimnospermas no ocurre la doble
fecundación, de manera que no hay endosperma verdadero. Las
semillas almacenan grasas, aceites y proteínas en el endosperma primario, protalo o
gametófito femenino haploide.
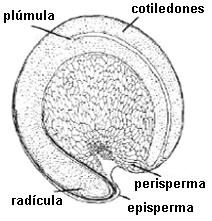
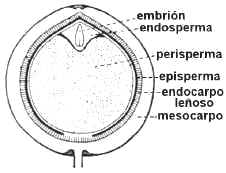
En Angiospermas hay tres posibilidades:
1.
Semillas albuminadas o endospermadas: se acumulan en el endosperma originado en la
doble fecundación. Los embriones presentan frecuentemente
cotiledones foliáceos o filiformes. Ejs.: Gramineae, Liliaceae, Palmae, Euphorbiaceae,
Umbelliferae.
En Myristicaceae, Annonaceae y Passifloraceae el endosperma es ruminado: excrecencias del episperma se
incrustan en el endosperma determinando que la superficie sea irregular.
2. Semillas perispermadas: el perisperma es el tejido
nucelar que perdura y se carga de sustancias de reserva. Se encuentra en Chenopodiaceae,
Amaranthaceae, Polygonaceae, y se considera como un carácter primitivo.
Algunas semillas tienen también endosperma, como
las de pimienta (Piper nigrum), Nymphaeaceae y Zingiberaceae.
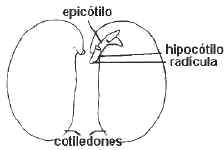
3. Semillas
exalbuminadas: Las sustancias de reserva se acumulan en los cotiledones. Ejs.:
nuez, muchas leguminosas: poroto, arveja y maní. Son las más evolucionadas, el
embrión tiene una plúmula con epicótilo desarrollado y varios primordios
foliares.
|
Fig. 6.89. Atriplex sp., semilla
perispermada
|
Fig. 6.89. Piper nigrum,
pimienta, corte long. de fruto. Semilla con endosperma y perisperma
|
fig. 6.90. Rosa carolina, semilla
exalbuminada en corte longitudinal de aquenio |
|
|
|
|
| Esquema de
Goebel (1933) |
Esquema de
Strasburger (1991) |
Esquema de
Robertson (1974) |
La más común es el almidón. Las semillas
con alto contenido en almidón tienen endosperma farináceo, como las de
gramíneas o poáceas.
Proteínas: se encuentran en una capa llamada aleurona
en los cereales, o como gluten, que determina las posibilidades de panificación
de las distintas harinas: capacidad de hacer masas consistentes y plásticas.
También se acumulan en los cotiledones, y tienen gran valor alimenticio, tanto
que reemplazan a las proteínas de origen animal: soja (Glycine max), arveja,
lenteja (Lens culinaris).
Grasas y aceites: Generalmente los lípidos se acumulan en los cotiledones como
en las nueces, el girasol y el maní, que son semillas oleaginosas.
Hemicelulosas:
se
acumulan en las paredes celulares, que se vuelven extremadamente gruesas, duras
y pesadas. El endosperma de las semillas de Phytelephas macrocarpa, palmera que
vive desde Panamá hasta el Perú, es muy duro; constituye el marfil vegetal.
Es
el conjunto de fenómenos por los cuales el embrión, que se halla en estado de
vida latente dentro de la semilla, reanuda su crecimiento y se desarrolla para
formar una plántula (plantita recién nacida).
Para que se produzca deben darse condiciones
fisiológicas entre las cuales las más importantes son oxigenación,
temperatura, luz (Lactuca sativa, Cecropia adenopus) y humedad: la absorción de
agua ocurre a nivel del hilo o la micrópila. El hinchamiento de la semilla
producido por la absorción de agua distiende los tegumentos seminales que
finalmente se rompen en la zona más débil, cerca de la micrópila.
Comportamiento
de las distintas partes del embrión
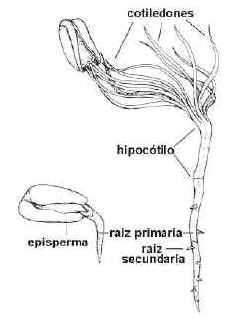
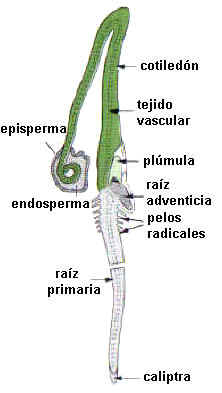
Radícula. Asoma por la micrópila,
dando origen a la raíz primaria. Su duración es efímera en las Monocotiledóneas que
generalmente desarrollan raíces adventicias, mientras en Gimnospermas y Dicotiledóneas
origina
la raíz principal que dura toda la vida de la planta.
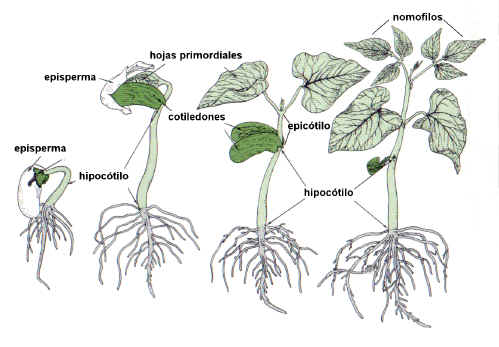
Hipocótilo. Su
crecimiento es importante en la germinación epígea, eleva los cotiledones por
encima del suelo. El episperma se rasga y los cotiledones, expuestos a la luz,
se vuelven los primeros órganos fotosintetizadores.
|
Fig. 6.91. Estados sucesivos de la
germinación epígea de
una semilla de Phaseolus, poroto.
|
 |
Dibujo de
Rost et al (1979) |
.
|
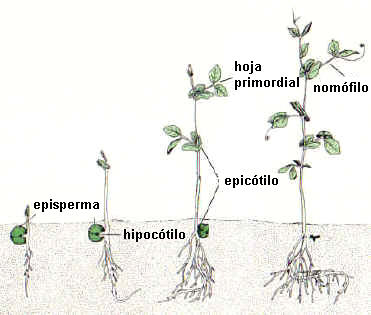
En la germinación hipógea su desarrollo es muy reducido o nulo,
los cotiledones quedan incluídos en el tegumento seminal por debajo de la
superficie del suelo. Ej.: arveja, Pisum sativum
|
Fig. 6.92. Estados sucesivos de la germinación hipógea
de una semilla de Pisum
sativum
|
|
|
| Cotiledones.
Su comportamiento varía según que se trate de semillas exalbuminadas o
albuminadas. En las semillas exalbuminadas se dan dos casos: si la germinación
es hipógea, como en Pisum, ceden las reservas acumuladas; si es epígea, como
en maní y poroto, ceden las reservas y luego enverdecen.
En las semillas
albuminadas también se dan dos casos. Si la germinación es hipógea cumplen
función haustorial como en Gramineae. Si la germinación es epígea, como en
Pinus, Ricinus, primero tienen función haustorial y luego enverdecen.
|
fig.
6.93. Estados
sucesivos de la germinación epígea de una semilla de Pinus,
pino
|
|
|
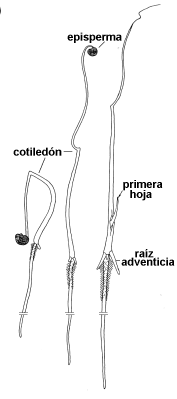
En Allium el único cotiledón crece alargándose durante la
germinación. Su porción apical permanece cubierta por el episperma y funciona
como haustorio, nutriéndose con el endosperma, mientras la parte basal queda
expuesta a la luz y es fotosintetizadora.
| fig.
6.94. Estados
sucesivos de la germinación de una semilla de Allium |
|
|
 |
|
Esquemas de Sachs |
Epicótilo. Tiene desarrollo precoz
solo en plántulas de germinación hipógea, como la arveja y muchas Monocotiledóneas,
elevando la plúmula por encima de la superficie del suelo. En
plantas de germinación epígea el epicótilo tiene desarrollo tardío.
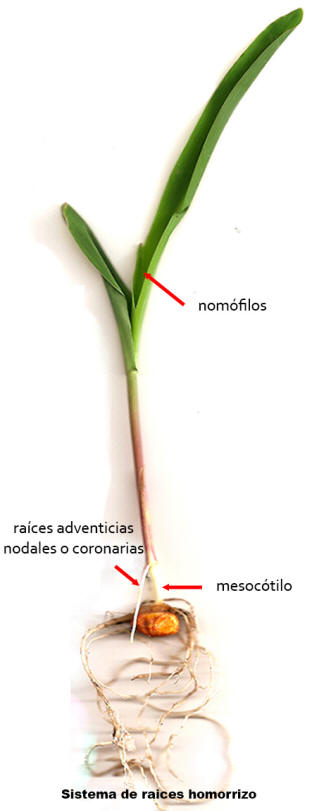
GERMINACIÓN DEL MAIZ
1. Puesto en
condiciones favorables a la germinación, el fruto de maíz se hincha como
consecuencia de la absorción de agua que genera un ablandamiento del
pericarpio y de los tejidos internos. En este momento la raíz primaria
adventicia rompe la coleorriza y sale al exterior creciendo varios
centímetros, inclusive se desarrollan raíces laterales a partir de ella.
Días posteriores, emergen numerosas raíces adventicias seminales
(1-13) que nacen del nudo cotiledonar que permanece dentro de la
semilla.
2. Se desarrolla el mesocótilo, porción del nudo cotiledonar que al
crecer separa el escutelo y la base del coleóptilo. Este
mesocótilo crece hacia la superficie a través de la tierra, llevando en el
extremo al coleóptilo que queda expuesto a la luz y fotosintetiza. El
coleóptilo protege a la plúmula y a varios primordios foliares en su
interior. El escutelo (como porción predominante del cotiledón) permanece
bajo tierra por lo cual la germinación es hipógea. El escutelo
funciona como haustorio, absorbiendo del endosperma las sustancias
nutricias y transfiriéndolas a las demás partes del embrión en desarrollo.
3. Más adelante, al desarrollarse la plúmula, el coleóptilo se rasga
permitiendo que asomen las primeras hojas, o nomófilos
fotosintetizantes.
4. Al poco tiempo, la raíz primaria adventicia junto a las raíces
adventicias seminales dejan de crecer, se secan lentamente y mueren.
Aparecen entonces nuevas raíces adventicias a nivel del nudo
cotiledonar o nudo 0 llamadas raíces adventicias
nodales o coronarias.
| fig. 6.96. Grano de maíz al
iniciar la germinación |
|
 |
|
Fig. 6.95. Estados sucesivos de la
germinación hipógea
de un grano de maíz |
|
 |
|
Fig. 6.95. Estados sucesivos de la
germinación hipógea
de un grano de maíz |
|
 |
Otro sistema de Clasificación de germinación
Según Duke (1969)
la germinación puede ser :
|
fanerocotilar, cuando los cotiledones emergen del
episperma y |
|
criptocotilar, cuando no
emergen del mismo. |
La fanerocotilar es más común en
Dicotiledóneas, y la criptocotilar en Monocotiledóneas. El maní es un tipo
transicional, y hay
muchos géneros que tienen los dos tipos en diferentes especies: Acer, Bauhinia, Caesalpinia,
Clematis, Couratari, Lecythis, Ormosia, Passiflora,
Phaseolus, Pithecellobium, Prunus, Quercus, Sapindus,
Sterculia, Terminales, Theobroma, etc. En Phaseolus la
criptocotilia es un carácter genético dominante sobre la fanerocotilia.
Según
Eames (1961) la germinación criptocotilar es más avanzada.
Semillas vivíparas: No tienen
dormancia, el
embrión simplemente crece fuera de la semilla y el fruto estando éste sobre la
planta madre. Ej.: Rhizophora mangle, mangle.
  
[ Tema 1 ] [ Tema 2 ] [ Tema 3 ] [ Tema 4 ] [ Tema 5 ] [ Tema 6 ] [ Tema 7 ] [ Tema 8 ] [ Tema 9 ] [ Tema 10 ] [ Tema 11 ] [ Tema 12 ] [ Tema 13 ] [ Tema 14 ] [ Tema 15 ] [ Tema 16 ] [ Tema 17 ] [ Tema 18 ] [ Tema 19 ] [ Tema 20 ] [ Tema 21 ] [ Tema 22 ] [ Tema 23 ] [ Actividades ] [ Tema 24 ]
|