|
|
|
Tema 24: Fecundación y embriogénesis
24.3. ANGIOSPERMAE
El proceso de la fecundación se inicia con la germinación
del grano de
polen sobre el estigma, desarrollando el tubo
polínico. La relación del tubo polínico con la intina no ha
sido totalmente aclarada: con microscopio óptico, el tubo
parece empujar a los lados la capa de intina que hay en la
abertura; con microscopio electrónico la intina se ve
continua con la pared del tubo. El tubo crece sobre el tejido
transmisor, sobre las células, entre ellas o en las paredes mismas. Las
paredes o las laminillas medias son disueltas por enzimas
pectinasas producidas en el extremo del tubo. El tubo
polínico crece por
alargamiento, esencialmente por un proceso de síntesis de
pared celular en la extremidad, llevado a cabo por dictiosomas
que aportan su contenido (sustancias pécticas y
hemicelulosas) para constituir la pared celular, mientras su
membrana alarga el plasmalema. |
Hacia arriba se forman tapones de calosa que sellan
sucesivamente las partes viejas vacías.
|
Germinación del polen y
desarrollo del tubo polínico en Angiospermas |
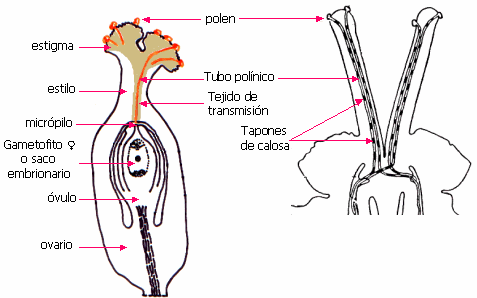 |
Muchos granos llegan al estigma y germinan, pero solo uno
producirá la fecundación de cada óvulo. El tubo sigue su
desarrollo sobre el tejido transmisor ovárico y llega al
óvulo, al que penetra generalmente por el micrópilo:
porogamia. Excepcionalmente puede penetrar por otros lugares:
aporogamia (Ulmus: tegumentos; Casuarina: cálaza). Los
óvulos de algunas plantas (Beta, Gasteria, Paspalum,
Ornithogalum) presentan un exudado micropilar viscoso que
contiene proteinas y carbohidratos. Posiblemente sirve como
fuente nutricional y también como guía para el tubo
polínico. En Gasteria el exudado parece ser secretado por el
aparato filar. (Franssen-Verheijen & Willemse, 1993).
En
algunas Gramineae (Andropogoneae y Paniceae: Stipa, Cenchrus,
Pennisetum, Brachiaria) las células del ápice de la nucela
se hipertrofian y proyectan al canal micropilar. Son células
de transferencia, tienen paredes gruesas, con invaginaciones y
protoplasma denso. Se ha sugerido que participan en la
nutrición del tubo polínico por su aspecto glandular. Esta
estructura se denomina embellum, término derivado del
inglés "embellishment" que significa ornamento
(Busri et al. 1993).
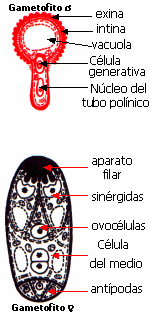
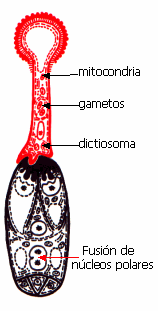
El citoplasma,
los gametos y el núcleo de la célula vegetativa se
encuentran en la porción apical del tubo polínico. Más
arriba se encuentra una gigantesca vacuola que aumenta de
tamaño por incorporación de agua.
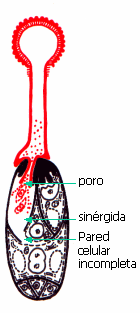
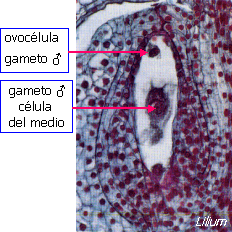
El tubo polínico hace contacto con el
saco embrionario en el aparato filar de la sinérgida, lo
atraviesa, y luego se forma un poro en el extremo del tubo para que pueda
descargar su contenido en el citoplasma de la sinérgida. Esta
recibe los gametos y parte de su citoplasma. El núcleo
vegetativo del tubo se desorganiza. Uno de los gametos penetra
en la ovocélula, y se fusiona con ella para constituir la
célula huevo o cigoto 2x. El otro penetra en la célula del
medio y se fusiona con el núcleo secundario formado por la
fusión de los dos núcleos polares, constituyendo el núcleo
primario del endosperma, generalmente triploide, 3x.
|
Este proceso,
llamado doble fecundación, es característico de las
Angiospermas. Desde la polinización hasta la fecundación
pueden transcurrir 12-48 horas (lo más frecuente).
Variantes: en
Taraxacum kok-saghys el proceso es muy rápido, requiere
apenas 15 minutos, mientras en Quercus dura 14 meses. |
Doble fecundación en
Lilium |
 |
|
Imagen de Rave |
La embriogénesis comienza generalmente
después que se inicia el endosperma, unas pocas horas
después de la fecundación, o después de varios meses.
El
cigoto muestra una diferenciación citológica entre los
polos calazal y micropilar, es decir que se
define un eje. La polarización del cigoto, proceso
controlado por el complejo membrana plasmática-citoesqueleto,
es la base del desarrollo ontogenético. El polo calazal es el
asiento de la mayor parte del crecimiento, el embrión
se desarrolla de células formadas en este polo. El polo
micropilar tiene esencialmente función vegetativa, su
crecimiento produce el suspensor que asegura al
embrión en la zona micropilar y suministra un mecanismo de
transferencia de nutrientes.
La primera división del cigoto en angiospermas es
transversal, excepto en las Piperaceae donde
aparentemente es longitudinal; las siguientes
pueden ser transversales, verticales u oblicuas. Los embriones
muestran una gran variación en la derivación de sus partes a
partir de los linajes de células producidas por las divisiones iniciales
en la cigota y sobre esta base se han establecido diversas
clasificaciones, muy complejas, con numerosas excepciones, que
no tienen mayor importancia porque se aplican al proembrión,
es decir a los primeros estadios del desarrollo, incluyendo el
suspensor.
Los estudios experimentales han permitido
constatar que no existe ningún patrón determinado de divisiones
celulares para originar los órganos del embrión (Rhagavan, 1997).