|
 |
|
TEMA 16
|
|
|
Tema 16.
TEJIDOS CONDUCTORES O VASCULARES
FLOEMA
|
El floema está íntimamente asociado al xilema,
formando el sistema vascular de la planta.
El floema es el tejido conductor encargado del
transporte de nutrientes orgánicos, especialmente azúcares,
producidos por la parte aérea fotosintética y autótrofa, hacia
las partes basales subterráneas, no fotosintéticas,
heterótrofas de las plantas vasculares.
El término floema fue acuñado por Nageli en
1858; deriva del griego "phloios" que significa corteza.
También se usan para designarlo los términos líber,
leptoma, tejido criboso.
Leptoma es un término debido a
Haberlandt y corresponde a las células del floema con paredes
primarias: elementos cribosos, células acompañantes y células
parenquimáticas; se le contrapone el término "hadroma"
para las células conductoras del xilema.
ORIGEN
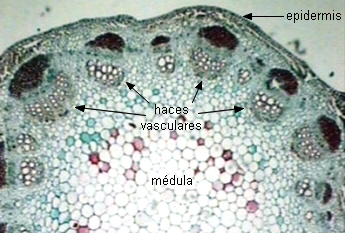
Se reconocen dos tipos de floema: el primario y el
secundario. En el vástago, el floema primario
se
encuentra asociado al xilema primario constituyendo los haces
vasculares.
| Sector de un transcorte de tallo primario
de Helianthus (girasol) |
|
|
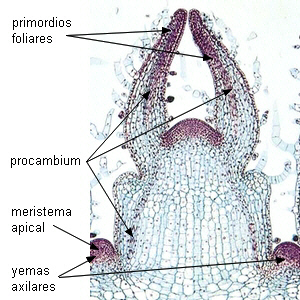
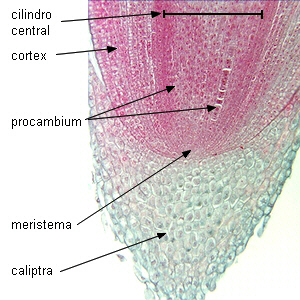
El floema primario, igual que el xilema primario,
se origina a partir del
procambium.
| Meristema apical caulinar:
Coleus |
Meristema apical radical: Zea
mays |
 |
 |
|
Imagen
de Mauseth |
Se diferencia en
protofloema y
metafloema. El
primero madura en las partes de la planta que aún están
creciendo en extensión, y sus elementos cribosos pronto se
vuelven inactivos. El
metafloema se
diferencia más tarde, completa su maduración después que el
órgano ha terminado su crecimiento en longitud. En las plantas
que no poseen crecimiento secundario, constituye el floema
funcional de los órganos adultos.
| Transcorte de
haces vasculares en Zea mays (Monocotiledónea) - Floema primario |
| Haz vascular joven |
Haz vascular maduro |
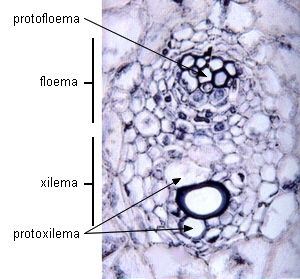 |
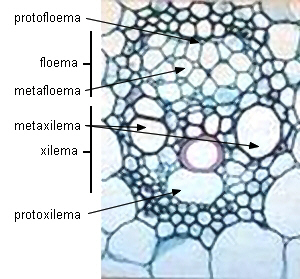 |
Imagen de Raven 2003 |
|
El
floema secundario, igual
que el xilema secundario, se
origina en el
cámbium, ubicado hacia la
periferia de tallo o raíz. Posee un sistema axial y un sistema
radial, que se continúa con el del xilema secundario a través del cámbium.
|
Sector de transcorte de tallo
secundario de Pinus
- Posición de los meristemas secundarios |
|
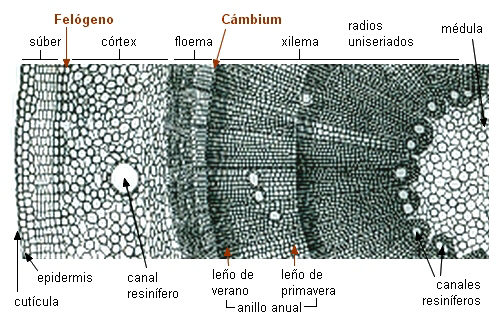
|
TIPOS DE CELULAS QUE COMPONEN EL FLOEMA
El floema, igual que el xilema, es un tejido complejo,
heterogéneo, formado por diferentes tipos de células: elementos
de conducción (elementos cribosos), elementos de sostén
(células esclerenquimáticas), elementos parenquimáticos y
elementos glandulares.
| Grupo |
Tipos de células |
Función |
| Elementos
cribosos |
Células
cribosas |
Conducción
de nutrientes orgánicos a larga distancia |
| Miembros de
tubos cribosos |
| Elementos
esclerenquimáticos |
Fibras |
Sostén, a
veces almacenamiento de azúcares |
| Esclereidas |
| Elementos
parenquimáticos |
Células
acompañantes |
Carga
y descarga de tubos cribosos: traslado radial
Almacenamiento
|
| Células
albuminosas |
| Células parenquimáticas axiales |
| Células parenquimáticas radiales |
| Elementos
glandulares e idioblastos |
Secreción,
depósito |
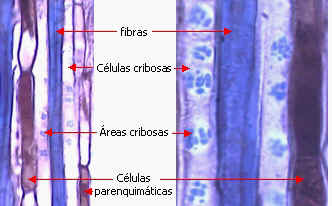
ELEMENTOS CRIBOSOS
Células del floema en Nicotiana tabacum,
en verde un tubo criboso |
|
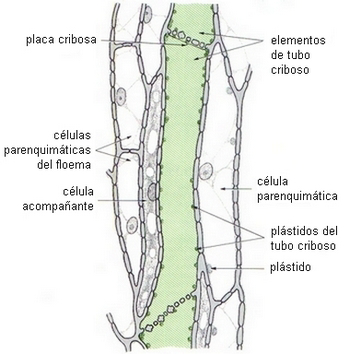
|
| Imagen de Rost, 1979 |
Son los elementos más especializados, se caracterizan por
sus protoplastos modificados y sus conexiones celulares
especiales. Fueron descubiertos por Hartig en 1837. Hay dos tipos:
células cribosas y
miembros de tubos cribosos. Sus
características comunes son:
 |
Pared celular |
Celulósica y primaria. Su espesor es variable,
algunas familias primitivas de angiospermas (Magnolia, Persea)
presentan paredes laterales con engrosamientos nacarados. Estos
espesamientos están compuestos por muchas capas de microfibrillas
de celulosa densamente dispuestas, y pectinas. En algunas especies
es tan marcado el engrosamiento que casi ocluye el lumen. Aparentemente
la función de estas paredes sería la de facilitar el trasporte radial
por apoplasto.
Algunas gimnospermas, según Esau, presentan engrosamientos
nacarados de naturaleza secundaria; otros autores no están de
acuerdo, consideran que se trata de paredes primarias.
|
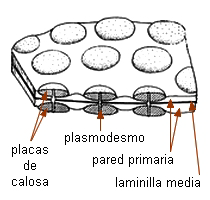
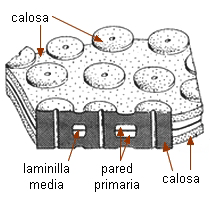
Comunicaciones intercelulares |
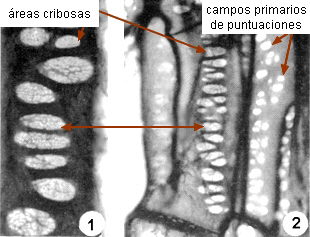
Los elementos cribosos se comunican entre
sí a través de áreas cribosas. Éstas son áreas
deprimidas de la pared provistas de poros a través de los cuales
se conectan los protoplastos de elementos vecinos por
medio de cordones citoplasmáticos. Se diferencian de los campos
primarios de puntuaciones por dos rasgos: 1) el tamaño de los poros,
generalmente mucho mayor que el de los plasmodesmos, se pueden
observar con
microscopio óptico, y 2) la presencia de un cilindro visible de
calosa, que rodea al cordón citoplasmático y puede aparecer
también en la superficie del área cribosa. |
Comunicaciones
intercelulares
en corte longitudinal de floema de Cucurbita sp. |
|
1-
áreas cribosas en paredes laterales de elementos cribosos
2-
campos primarios de puntuaciones en células
parenquimáticas
|
|
|
|
Imágenes
de Esau 1953 |
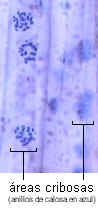
La calosa es un carbohidrato, polímero de la glucosa con enlaces
ß1-3. Es sintetizada en la membrana plasmática, aparentemente
por las mismas rosetas de proteínas enzimáticas que sintetizan la
celulosa. No es refringente, se tiñe de azul con azul de anilina
o azul de resorcina. Se encuentra también una delgada capa de
calosa rodeando cada plasmodesmo, está en los tubos polínicos, rodeando
las células madres de los granos de polen, en cistolitos y en las
paredes celulares de ciertos hongos.
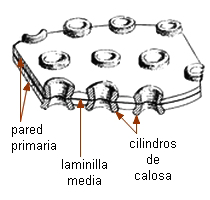
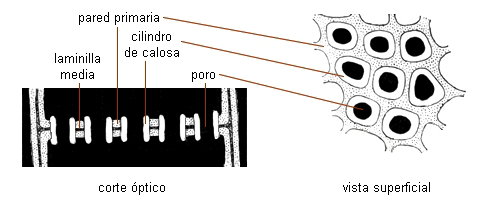
Las áreas cribosas vistas de frente se ven como una zona
deprimida con un número variable de poros rodeados por anillos de
calosa. En corte se observan como porciones adelgazadas de la
pared, con cordones que pasan de una célula a la otra.
Las áreas cribosas son menos especializadas: sus poros y cordones
citoplasmáticos son de menor diámetro, igual que los cilindros
de calosa. Se hallan generalmente en las paredes laterales de los
elementos cribosos.
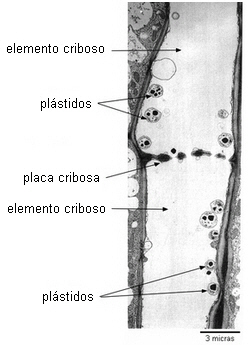
Las placas cribosas son más especializadas, sus poros son de gran
diámetro, hasta 15 µ, y generalmente se encuentran en las
paredes terminales de los elementos de tubos cribosos.
| Esquema de una placa
cribosa |
|
|
|
Imagen de Esau
1972. |
Hay dos tipos de placa cribosa: simple y compuesta.
| La placa cribosa simple
consta de una sola área cribosa. |
Placa cribosa simple en
transcorte de tubo criboso de Cucurbita
sp., zapallo
|
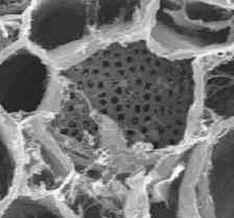 |
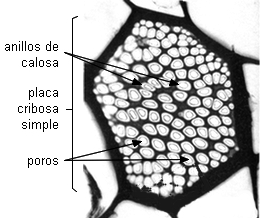 |
Foto MEB,
Ana Ma. Gonzalez |
Imagen de Esau
1953 |
Las placas cribosas se encuentran generalmente en las paredes
terminales casi horizontales de los tubos cribosos.
|
Placa cribosa simple en Cucumis |
| en corte longitudinal |
en corte transversal |
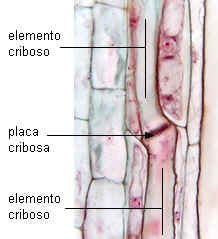 |
 |
|
Imágenes de Mauseth, |
Las placas cribosas compuestas presentan varias a numerosas áreas
cribosas.
| Placas
cribosas compuestas en miembros de tubos cribosos - corte
longitudinal. |
| Esquema en Nicotiana tabacum |
Vista superficial en
corte radial de Salix sp |
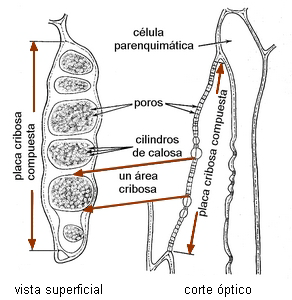 |
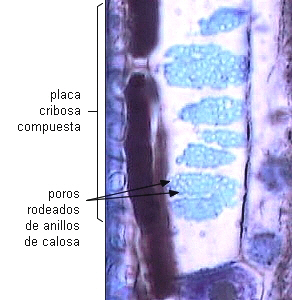 |
|
Esquemas de Esau 1972 |
|
|
Formación de poros y deposición de calosa |
Se ha
comprobado que la calosa se sintetiza rápidamente como respuesta
a injurias. Se encuentra también en suspensión en el citoplasma
y cuando la célula es lastimada precipita contribuyendo a taponar
los poros.
En plantas superiores, el material procesado con
técnicas para microscopía óptica y electrónica muestra acumulaciones características de calosa en los futuros poros
de las células en diferenciación; en cambio en algunas plantas inferiores no aparece calosa en el desarrollo
de los poros (Esau & Thorsch 1985). En
Lemna minor no se deposita calosa durante la formación de los
poros (Walsh & Melaragno 1976).
|
Secuencia
de formación de poros - Esquemas de ultraestructura
|
|
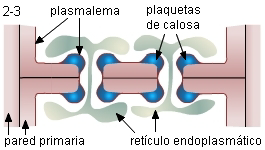
1. El sitio de la pared donde se formará cada poro está
atravesado por un plasmodesmo, y frecuentemente en ese lugar la
pared es más gruesa. Cisternas circulares del RE, próximas a la
pared se hallan asociadas al plasmodesmo en ambas células, y
conectadas a través del desmotúbulo. Determinan así la zona de
deposición de calosa.
|
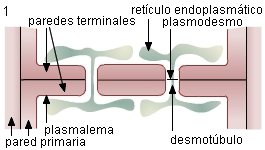 |
2. La calosa se deposita en forma de plaquetas entre
la membrana plasmática y la pared primaria. Las plaquetas incrementan gradualmente en
superficie y espesor.
3. La pared celular aumenta de espesor, especialmente entre los
futuros poros. A la altura de los futuros poros aparentemente la
fase amorfa de la pared primaria y finalmente la laminilla media
son reemplazadas por calosa. |
 |
|
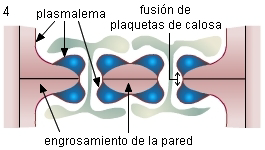
4. Se produce la fusión de las plaquetas de las dos células
formando el cilindro de calosa. La masa de calosa que se acumula
se corresponde en tamaño con el lumen del futuro poro.
|
 |
|
5. Las cisternas del RE se fraccionan en una serie de vesículas
que se apartan del poro, el desmotúbulo se desorganiza, y la
manga citoplasmática que lo rodeaba se transforma en el cordón
citoplasmático. La membrana plasmática permanece rodeando los
cordones. Por último, la mayor parte de la calosa se disuelve,
formándose el lumen del poro (Esau & Thorsch 1985) |
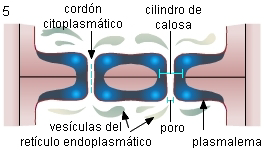 |
A medida que el elemento criboso envejece, aumenta la cantidad de
calosa en los cilindros, el diámetro de los poros disminuye y se
comprimen los cordones citoplasmáticos. Por último la calosa
puede depositarse sobre toda la superficie del área cribosa, que
de ser una depresión pasa a ser una zona engrosada. La
calosa forma una verdadera almohadilla.
En otoño, en algunas especies como la vid, Vitis, las áreas cribosas son
bloqueadas por estas masas de calosa, deteniendo el flujo de la
savia. En primavera el citoplasma disuelve la calosa por acción
enzimática, y se renueva el pasaje de savia.
Flujo estacional de la
calosa en tubos cribosos de Vitis |
|
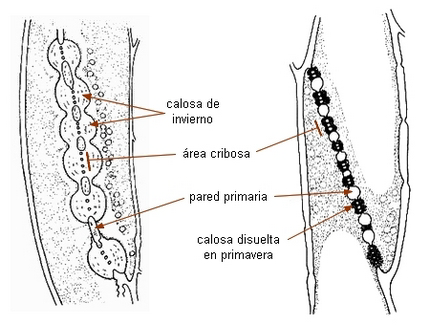
|
|
Imágenes de Fahn 1990 |
Cuando un elemento criboso cesa definitivamente en su actividad,
se forma la calosa definitiva, llamada "callo" por autores
antiguos, el protoplasto se desorganiza y desaparecen los cordones
de conexión.
|
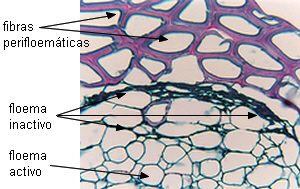
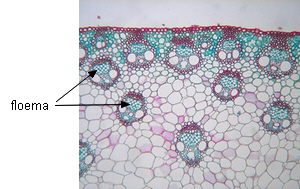
Longevidad |
En la mayoría de las dicotiledóneas los elementos
cribosos son funcionales sólo durante una estación vegetativa, a
veces sólo días o semanas, siendo reemplazados por otros nuevos.
En algunas plantas leñosas pueden durar varios años. En las
palmeras los elementos del metafloema duran toda la vida de la
planta.
| Dicotiledóneas (floema
estacional) |
Monocotiledóneas (floema
permanente) |
 |
 |
| Imagen de Mauseth |
Transcorte de tallo de Zea
mays |
Diferenciación de los elementos cribosos
A la madurez los elementos cribosos son células vivas, pero durante
su ontogenia el
protoplasto experimenta cambios profundos. Los rasgos
característicos en la diferenciación de los elementos cribosos,
son:
NÚCLEO: comúnmente ocurre la degeneración del núcleo;
en Angiospermas se vuelve
menos coloreable - degeneración cromatolítica, mientras en Gimnospermas
se vuelve muy oscuro y arrugado - degeneración picnolítica (Fahn 1990). En
algunas Angiospermas se conserva: Vitis, Robinia,
Ulmus. También pueden conservarlo las células cribosas de
algunas Gimnospermas: Taxodiaceae (Sequoia, Metasequoia,
Taxodium). CITOPLASMA: la membrana
plasmática, límite externo del citoplasma, se conserva. En
cambio el tonoplasto, que la separa de la vacuola, se desorganiza,
de modo que el límite entre citoplasma y vacuola
desaparece. La mezcla recibe el nombre de mictoplasma.
Las cisternas del retículo endoplasmático (RE) se localizan en los
sitios donde se formará la placa cribosa y ocurrirá la
deposición de calosa. Después se reorganiza, se vuelve liso y la
mayor parte se agrupa en pilas.
Los dictiosomas se encuentran activos durante el engrosamiento de la pared. Se
degeneran junto con el núcleo, igual que los ribosomas y el
citoesqueleto. Los únicos orgánulos celulares que se conservan
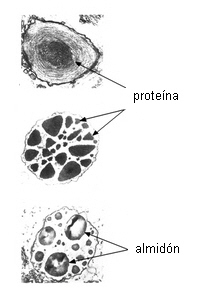
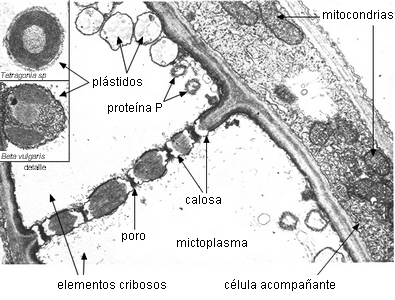
son las mitocondrias y los plástidos. PLASTIDOS.
Hay dos tipos de plástidos en los elementos cribosos:
P y S (Behnke, 1981). Los plástidos P son los que tienen depósitos de proteína
acompañados o no de almidón. Los plástidos S son los que
presentan almidón exclusivamente, de forma, tamaño y estructura
variable. En los elementos adultos los plástidos pueden carecer
de proteina o almidón, pero durante la ontogenia se observan en
los mismos acumulaciones de uno u otro compuesto.
Las Gimnospermas presentan plástidos S, excepto las Pinaceae con
plástidos P. En las Angiospermas ell tipo principal es
P, con inclusiones cristaloides o filiformes. Todas
las Monocotiledóneas presentan un subtipo característico
ultraestructuralmente distinto de plástido P. Las Dicotiledóneas
presentan ambos tipos; hay ciertos grupos, como las
Centrospermales y las Leguminosas que también poseen subtipos
característicos de plástidos P. En cambio, determinados grupos de
dicotiledóneas que en otros aspectos se consideran avanzadas,
como las Asterideae, Dilleniidae y la mayoría de las
Rosideae,
presentan plástidos de tipo S (Behnke 1981)
|
Plástidos
en los elementos cribosos
Placa cribosa madura en floema de Nicotiana tabacum
|
|
|
|
Imagen de Esau 1977 |
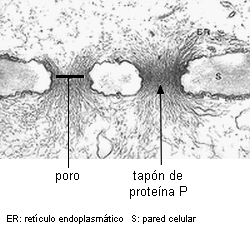
PROTEINA P. Es una de las características más importantes de los
tubos cribosos de Dicotiledóneas, es raro en tubos cribosos
de Monocotiledóneas y está ausente en las células cribosas de
Gimnospermas (Fahn 1990).
Se sintetiza en los tubos jóvenes nucleados, y se acumula en los
cuerpos de proteina P o cuerpos mucilaginosos, de tamaño
variable, compuestos de túbulos o filamentos de proteina
apretadamente dispuestos. En los tubos maduros, enucleados, la
proteina P se distribuye en todo el lumen en la mayoría de los
casos. Cuando adopta forma fibrilar, los filamentos están constituidos por
dos subunidades dispuestas helicoidalmente en posición parietal y
en los poros de la placa cribosa. Si la célula es injuriada, se
encuentra en todo el lumen celular, y obstruyendo los poros de las
áreas cribosas, produciendo la formación de un tapón
mucilaginoso que detiene la exudación de savia en los primeros
momentos.
Proteína P
en los poros de una placa cribosa
en Aristolochia brasiliensis |
Proteína P en corte
longitudinal de
floema en Cucumis |
|
|
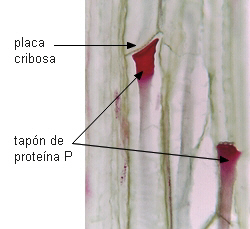
|
| Fotomicrografía
electrónica de Behnke en Bergfeld et al.
|
Imagen de
Mauseth |
En muchas familias de dicotiledóneas leñosas se forman cuerpos
de estructura cristalina que no se dispersan, y persisten en los
tubos enucleados (antiguamente algunos fueron descriptos como
"nucleos extruidos"); según su forma se clasifican en
cuatro grupos: compuestos esféricos, globulares, fusiformes,
estrellados; cada célula contiene un solo cuerpo, de tamaño
variable.
Las monocotiledóneas no presentan cuerpos cristalinos, y también
están ausentes en muchas familias herbáceas de dicotiledóneas.
Transcorte de floema de
Eucalyptus sp
esta imagen no se ve con el explorador |
|
|
|
Imagen de Esau 1972 |
CELULAS CRIBOSAS
Se encuentran en Pteridofitas y
Gimnospermas. Se comunican entre sí por áreas cribosas,
que están dispersas en toda la superficie de la célula.
En las Pteridofitas las células cribosas son largas,
aguzadas y enucleadas, con áreas cribosas pobremente
diferenciadas. Usualmente tienen esférulas, cuerpos
proteicos limitados por una membrana.
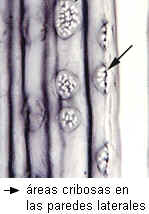
Las células cribosas de las Gimnospermas son elementos largos y
delgados, con extremos afilados, que se superponen. En Sequoia
las áreas cribosas se encuentran sobre las paredes radiales.
| Sequoia: células
cribosas en corte longitudinal radial |
Pinus |
|
|
|
|
|
|
Imagen de
Mauseth |
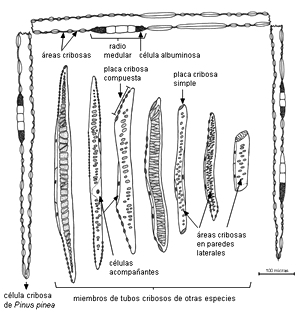
MIEMBROS o ELEMENTOS DE LOS TUBOS CRIBOSOS
Se encuentran en Angiospermas.
Son series longitudinales de células llamadas 'miembros de tubos cribosos'
conectadas entre sí por medio de placas cribosas simples o
compuestas. En las paredes
laterales tienen áreas cribosas más o menos especializadas,
generalmente difíciles de ver.
Cyathea gigantea y varias especies de Equisetum constituyen una
excepción entre las Pteridophyta, ya que poseen tubos cribosos con placas cribosas.
Tipos de elementos
cribosos en plantas con semilla |
|
|
|
Imagen de Esau 1977 |
Diferencias entre células cribosas y miembros de tubos cribosos
|
Célula cribosa |
Miembro de tubo
criboso |
| Célula larga y
delgada con paredes terminales aguzadas |
Célula más corta
y ancha con terminales inclinadas u horizontales |
| Areas
cribosas poco especializadas en paredes laterales y
terminales |
Placas cribosas en
paredes terminales |
| Áreas cribosas en
paredes laterales |
| Sin proteína P |
Con
o sin proteína P |
| Asociada con
células albuminosas morfológica y fisiológicamente |
Asociado con
células acompañantes ontogénica, morfológica y
fisiológicamente |
| Pteridophyta -
Gimnospermae |
Angiospermae
excepto Austrobaileya scandens y
Sorbus aucuparia. Pteridophyta: Equisetum, Cyathea
gigantea |
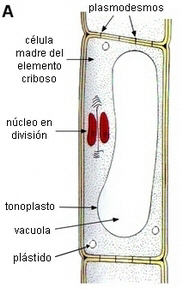
ONTOGENIA DE UN ELEMENTO CRIBOSO
En las cuatro figuras siguientes se esquematizan
los diferentes estadios de la ontogenia:
|
Secuencia de
formación de un elemento criboso |
| A.
Célula precursora de un elemento criboso, con leucoplastos y
núcleo en división.
|
 |
| B.
Como resultado de la división se forman dos células: un
elemento criboso con paredes gruesas en desarrollo,
núcleo conspicuo, grande, vacuola, cuerpos de proteína
P, plástidos, y plasmodesmos en las paredes
terminales. La segunda célula, es la célula
acompañante.
|
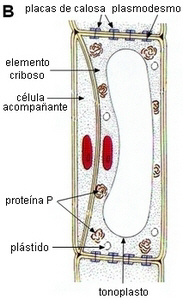 |
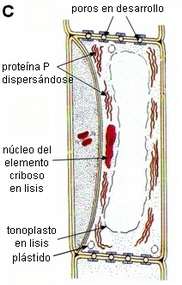
| C.
El elemento criboso muestra el núcleo degenerado,
el tonoplasto parcialmente destruido, la proteína P dispersa,
los plástidos ubicados en el citoplasma periférico;
en los
futuros poros de la placa cribosa se observan plaquetas de calosa depositadas.
En muchas especies la célula acompañante se divide en dos
o más células.
|
 |
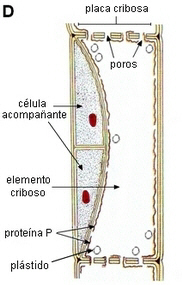
| D.
El elemento criboso maduro muestra los poros abiertos en
la placa cribosa,
bordeados de calosa, y la proteína P dispersa en el citoplasma
periférico con RE y
plástidos, y una parte central que es una mezcla de jugo celular
y material citoplasmático desorganizado: el mictoplasma.
|
 |
|
Imágenes modificadas de Raven
2003 |
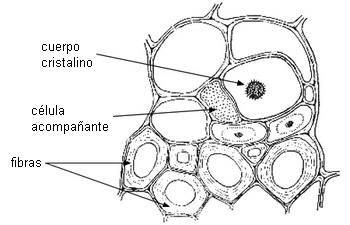
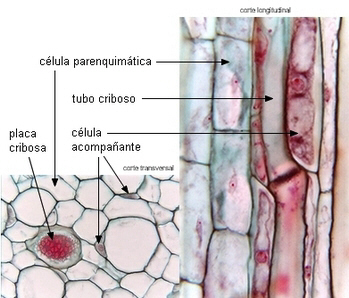
CÉLULAS ACOMPAÑANTES
Son células parenquimáticas muy especializadas, asociadas
ontogenéticamente con los miembros de los tubos cribosos en el
metafloema y floema secundario de Angiospermas. Algunas están
diferenciadas como células de transferencia.
|
Células acompañantes
de tubos cribosos de Angiospermas |
| Transcorte
de Cucumis sp. |
Corte
longitudinal de Asclepias |
|
|
|
Imágenes de Mauseth |
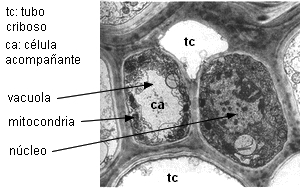
Tienen pared primaria con campos primarios de puntuaciones con
plasmodesmos ramificados, enfrentados a los poros de las áreas
cribosas de los elementos cribosos. Durante la ontogenia, se
deposita calosa del lado del elemento criboso, pero no del lado
de la célula acompañante, donde permanencen los campos
primarios de puntuaciones (Esau & Thorsch 1985).
|
Detalle de la comunicación intercelular
mixta
entre una célula acompañante y un miembro de tubo criboso
|
|
|
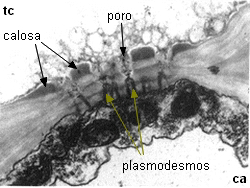
|
| Imágenes
de Mauseth 1991 |
Su protoplasto es el característico de las células
metabólicamente activas: con núcleo grande frecuentemente
poliploide, nucléolos grandes, vacuolas pequeñas, RE bien
desarrollado, grandes mitocondrias, dictiosomas, abundantes
ribosomas. Pueden tener cloroplastos y leucoplastos, pero no
forman almidón. Asumen las funciones nucleares de los elementos
cribosos, mueren cuando éstos dejan de ser funcionales. Cumplen
la función de carga y descarga de los elementos cribosos,
trasportando lateralmente los fotosintatos (Fahn 1990).
Localización
Puede no haber células acompañantes en el
protofloema de Angiospermas (Esau 77). En las Gramineae se halla
una disposición muy regular de tubos cribosos y células
acompañantes. Se ha comprobado que esta disposición está
correlacionada con tipos de haces vasculares avanzados, mientras
que la disposición irregular ocurre en tipos más primitivos de
haces.
|
Floema primario en transcorte de haz
vascular de Zea mays
|
|
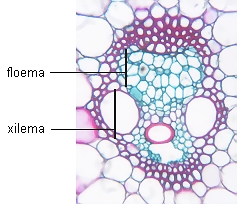
|
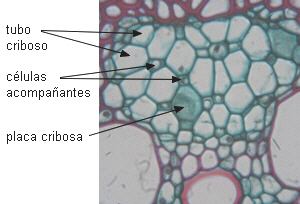
|
Ontogenia
Se forman a partir de la misma célula meristemática
que los miembros de los tubos cribosos. Esta se divide
longitudinalmente una o más veces, dando células de diferente
tamaño. La célula mayor se diferenciará en miembro del tubo
criboso, y la célula restante formará las células
acompañantes previa división transversal que puede no ocurrir.
En resumen, un miembro de tubo criboso puede tener asociado un
número variable de células acompañantes, dispuestas en series
longitudinales.
CELULAS ALBUMINOSAS o de Strasburger
Son células del parénquima radial o vertical del floema
secundario de gimnospermas. Se tiñen intensamente con colorantes para sustancias proteicas.
Tienen comunicaciones intercelulares con las células cribosas y
nunca contienen almidón. Presentan alta actividad enzimática y
respiratoria asociada con la actividad de carga y descarga de
las células cribosas (Fahn 1990). Cumplen
igual función que las células acompañantes, pero tienen un
origen ontogenético diferente: no se forman a partir de la
misma célula meristemática (excepto en Ephedra, Fahn 1990). La
asociación es morfológica y funcional, semejante a la antes
descripta. Nunca han sido descriptas paredes laberínticas en
estas células.
| Esquema |
Corte longitudinal
tangencial de floema de Pinus strobus |
|
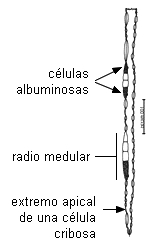
|
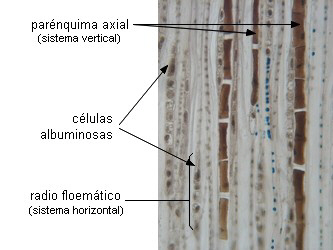
|
CELULAS PARENQUIMATICAS
Existen en cantidad variable, y son menos especializadas que las
células acompañantes o las células albuminosas. En el floema primario son
alargadas paralelamente a los tubos; en el floema secundario se
presentan en el sistema vertical y en el horizontal. En el
vertical están en dos formas básicas: células fusiformes o
hileras de células. En el horizontal constituyen los radios del
floema, integrados por dos tipos de células: procumbentes,
alargadas en dirección radial y erectas, generalmente
marginales, alargadas en sentido vertical.
Pueden estar diferenciadas en células de transferencia, con
paredes laberínticas.
|
Células
|
|
Sistema
Vertical |
Sistema
Horizonal
|
Sequoia
sempervirens:
células |
Tilia
americana
célula |
Salix sp.:
célula
con
taninos |
procumbentes
(p) y erectas (e)
en
radio floemático de Salix
|
|

|
|

|
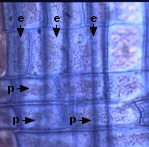
|
Funciones
Participan en la carga y descarga de los elementos
cribosos trasportando azúcares a las células acompañantes.
Almacenan almidón, grasas, taninos y cristales.
CÉLULAS INTERMEDIARIAS
Son células parenquimáticas y células acompañantes, en el
floema de las venas venores de las hojas, que sirven como
conección entre el tejido fotosintético y el sistema de
elementos cribosos. Frecuentemente están diferenciadas en
células de transferencia en las
Angiospermas. En las
gimnospermas hasta ahora no han sido encontradas paredes
laberínticas, pero como en muchas angiospermas las células
intermediarias pueden tener paredes lisas, se piensa que las
células albuminosas son también células intermediarias
(Mauseth 1989).
ELEMENTOS ESCLERENQUIMÁTICOS
|
Fibras
|
En el floema primario pueden ser muy largas, se disponen
externamente. En el floema secundario tienen distribución
variable, en bandas o dispersas, y son más cortas. Depositan
paredes secundarias, y especialmente las del floema secundario,
se lignifican. También hay fibras septadas.
|
Corte transversal de
estructura primaria de tallo |
|
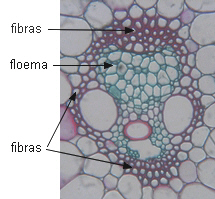
|
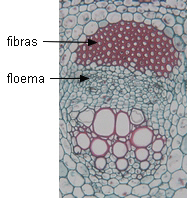
|
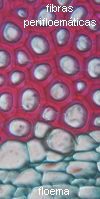
|
| vaina de fibras en Zea
mays |
fibras en
posición periférica en Helianthus sp |
Floema
en transcorte de tallo
secundario |
|
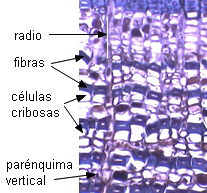
|

|
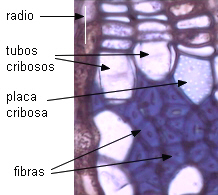
|
|
bandas
de fibras en Sequoia
sempervirens
|
grupos
de fibras en Tilia americana |
|
Esclereidas
|
Las esclereidas del floema secundario aparecen
generalmente por esclerificación de células
parenquimáticas. Pueden presentarse solas o estar combinadas
con fibras.
Nidos
de braquiesclereidas en
floema secundario de Quercus alba
|
 |
 |
|
corte
longitunidal tangencial |
detalle |
ESPECIALIZACION FILOGENETICA DEL FLOEMA
No hay datos suficientes sobre la
especialización filogenética del floema porque el tejido generalmente
no se conserva en los fósiles, y aún en plantas vivientes requiere técnicas
especiales de investigación. Su estructura varía con el
manipuleo por su sensibilidad y capacidad de reacción ante las lesiones.
Tendencias evolutivas observadas en miembros de tubos cribosos
1. Desarrollo de placas cribosas en paredes terminales.
2. Cambio de orientación de pared terminal, de oblicua a
transversal.
3. Cambio de placas cribosas compuestas a placas simples, y
disminución del número de áreas en las paredes laterales.
Comparación con los elementos traqueales del xilema
| Xilema |
Floema |
| Traqueida (extremos superpuestos con puntuaciones) |
Célula cribosa (extremos superpuestos con áreas cribosas) |
| Miembro de vaso (placa de perforación) |
Miembro de tubo criboso (placa cribosa) |
| Perforación: desaparición de la pared primaria |
Poros:
más amplios en la placa cribosa |
| Placa
de perforación:
de escalariforme o foraminada a simple |
Placa cribosa:
de compuesta a simple |
| Orientación pared terminal
: oblicua a transversal |
Orientación pared terminal
: oblicua a transversal |
|
|